Warning: replacing previous import 'S4Arrays::makeNindexFromArrayViewport' by
'DelayedArray::makeNindexFromArrayViewport' when loading 'SummarizedExperiment'
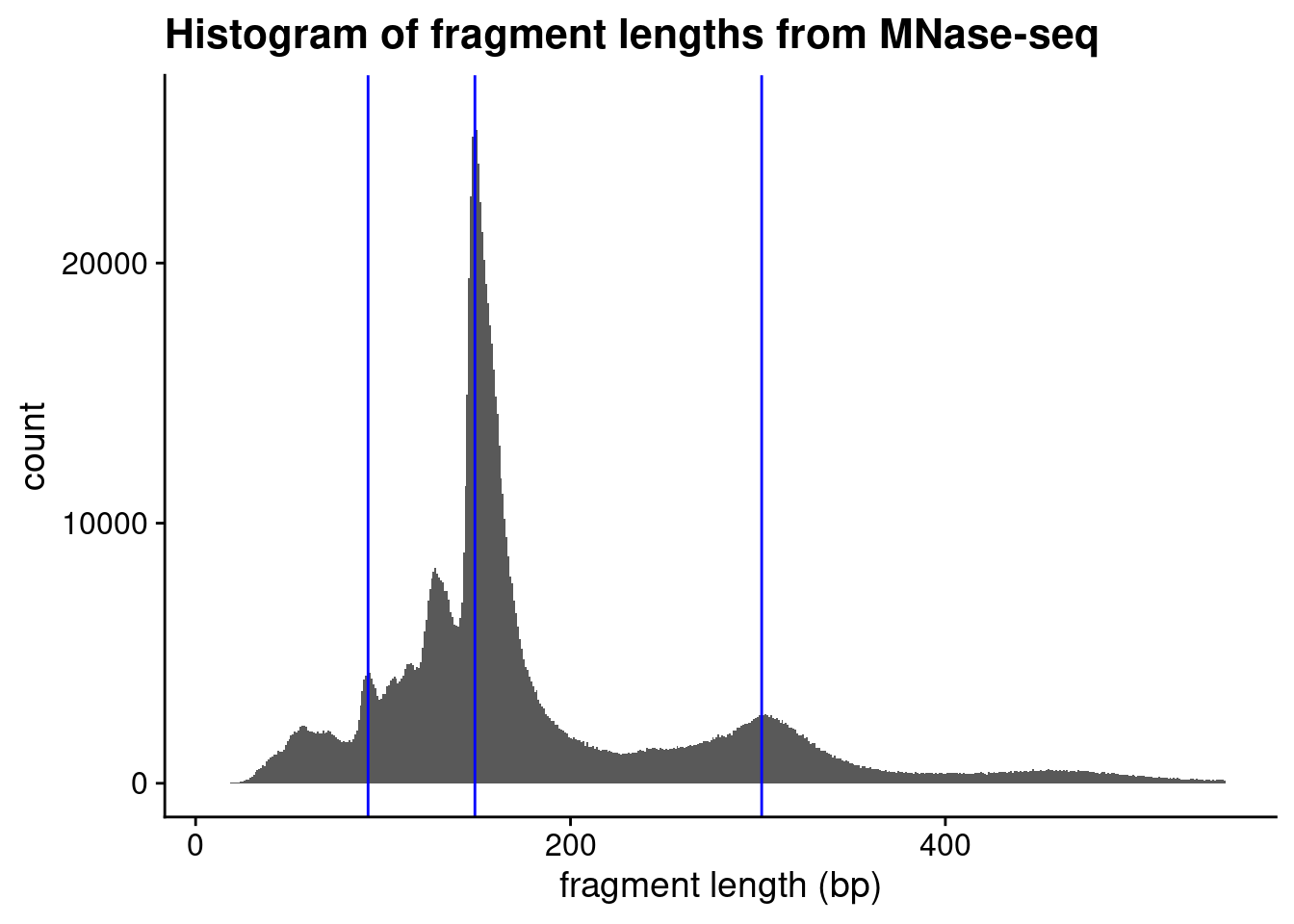
In class we learned that MNase digestion yields nucleosomal “footprints” of ~150 bp in size. I’ve added blue vertical lines to emphasize positions of the major peak (intact nucleosomes) as well as smaller “sub-nucleosomal” peak.
We can calculate the counts for the histogram above and more precisely determine the maximum signal using which.max() to identify the index of the maximum value in a vector (not the value itself!):
frag_hist<-mnase_tbl|>mutate(frag_len =end-start)|>count(frag_len)# `which.max` takes a vector and gives us the index of the maximum valuemax_idx<-which.max(frag_hist$n)# now we can use index to find the abundant fragment sizencp_max<-frag_hist$frag_len[max_idx]
So this tells us that that the most abundant fragment size in the library is 149 bp.
Question 1 – 7 points
Let’s take a closer look at the some of the smaller fragments in the MNase experiment. In particular, let’s zoom in on the populations of fragments that are smaller than 1 nucleosome in size, the peak between 85 and 95 bp (the left-most blue vertical line).
Use the above strategy to precisely determine the peak of this smaller size range. How big are those fragments? These are called “sub-nucleosomal” fragments.
Recreate the histogram using ggplot2 (using relevant code from class 17) and add the blue vertical lines at the peak positions you calculated, including the position of the disomes above.
ggplot(mnase_tbl,aes(x =end-start))+geom_histogram(# single base-pair resolution binwidth =1)+labs( x ="fragment length (bp)", title ="Histogram of fragment lengths from MNase-seq")+theme_cowplot()+geom_vline(xintercept =c(subnuc_max, ncp_max, dinuc_max), color ="blue")
Question 2 – 13 points
Next we’re going to look at where these sub-sucleosomes are with respect to intact nucleosomes.
Our strategy will be to compare the density of sub-nucleosomes relative to the mid-points of previously mapped nucleosomes. Specifically, our reference point will be the midpoints of the +1 nucleosomes.
So you’ll make a metaplot, but instead of using transcription start sites as the reference point, we’ll use the midpoints of the +1 nucleosome, and instead of MNase-seq signal density, you’ll count up the number of individual reads that intersect with windows around those midpoints.
First, load the relevant data. We’ll re-use the yeast_mnase_chrII.bed.gz data you loaded above, plus you’ll need to load two other files:
a “genome” file, sacCer3.chrom.sizes
a BED file, yeast_p1_chrII.bed.gz which contains the mid-points of the +1 nucleosomes on chromosome 22. Recall that the +1 nucleosome is the nucleosome downstream of the transcription start site.
Next, we need the mid-points of nucleosomes for comparison. The following function needs fixing.
calc_mids<-function(tbl, min_len, max_len){tbl|>mutate(# XXX: blanks here frag_len =end-start)|>filter(# XXX: blanks herefrag_len>=min_len&frag_len<=max_len)|>mutate(# XXX: blanks here midpoint =start+round((end-start)/2))|>select(chrom, midpoint)|>rename(start =midpoint)|>mutate(end =start+1)}
ncp_mids_tbl<-calc_mids(mnase_tbl, ncp_max-3, ncp_max+3)|>bed_slop(genome, both =20)subnuc_mids_tbl<-calc_mids(mnase_tbl, subnuc_max-3, subnuc_max+3)|>bed_slop(genome, both =20)
Next, we need to make the intervals for the metaplot. We’ll look 100 bp up- and downstream of the +1 nucleosome positions, and make windows that are 1 bp in size.
p1_win_tbl<-bed_slop(p1_tbl,genome, both =100)|># XXX: blank herebed_makewindows(win_size =1)
Almost there! Now you just need to identify the number of short and long nuclesome fragments (based on their midpoints) that intersect with the +1 nucleosomes you defined above.
Use bed_intersect() to identify fragments that overlap, and then just count the number of fragments per .win_id (don’t forget the suffix). Note you will do this separately for the short and long fragments.
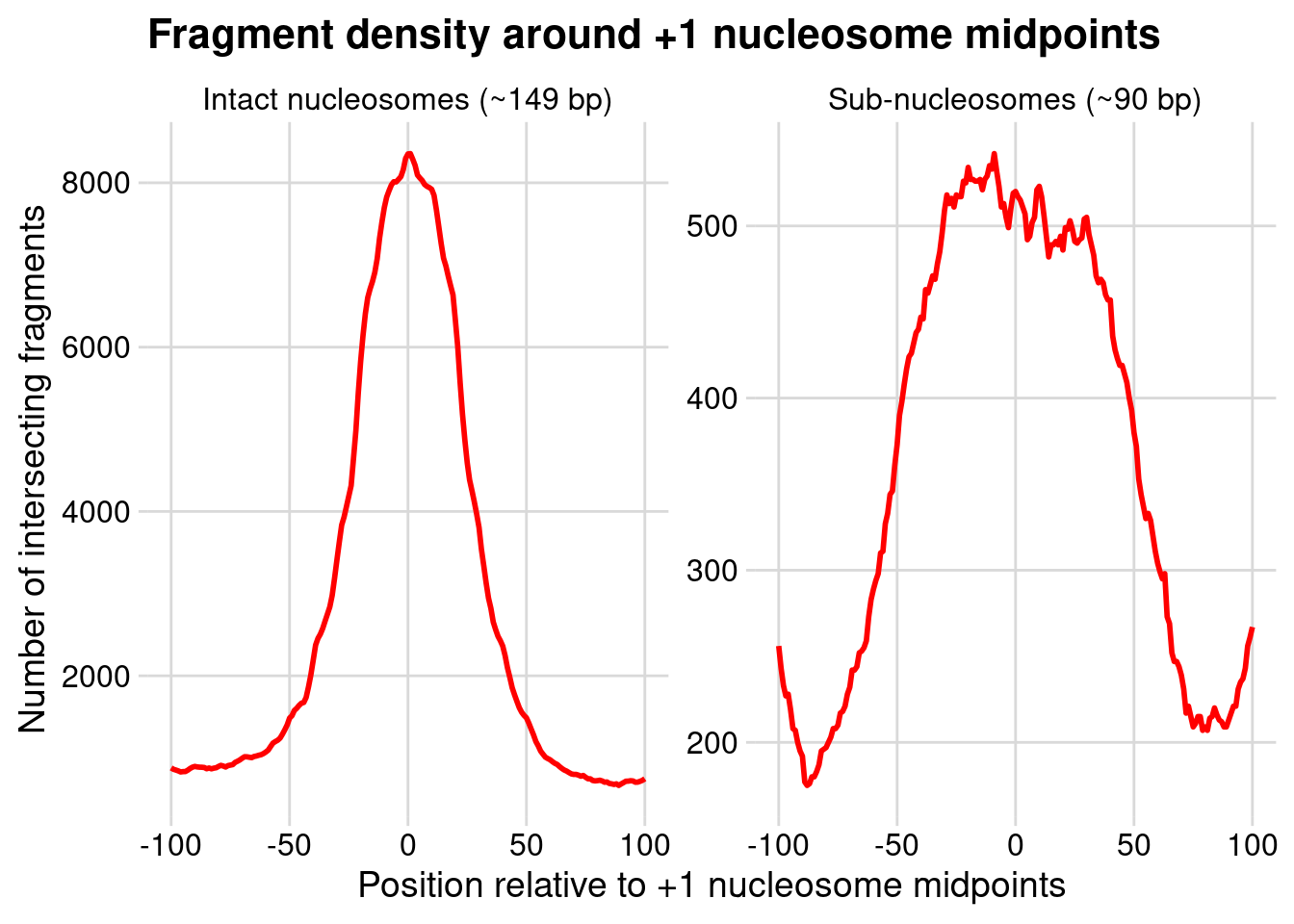
Finally, we plot the data with position on the x-axis, and count on the y-axis.
ggplot(all_tbl,aes(win_ids, n))+geom_line( linewidth =1, color ="red")+facet_wrap(~type, scales ="free_y")+theme_minimal_grid()+labs( x ="Position relative to +1 nucleosome midpoints", y ="Number of intersecting fragments", title ="Fragment density around +1 nucleosome midpoints")
Interpretation
How do you interpret these plots?
Rationalize the pattern for intact nucleosomes. What pattern did you expect to see?
Expectation is that large nucleosomal fragments will be centered on +1 nucleosome midpoints, so the plot agrees. The width of the density has to do with the size of the intervals we defined (expanding single-base pair midpoints by 20 and 100 bp (for query and reference intervals, respectively).
Rationalize the pattern for sub-nucleosome. How would you describe the positions of sub-nucleosomal fragments, relative to the +1 nucleosome midpoints? What might this mean with respect to gene transcription?
The density is bimodal, indicating two distributions of sub-nucleosomal fragments relative to +1 nucleosome midpoints. These fragments are formed at the edges of the nucleosome core particl during the process of nucleosome disassembly during transcription (losing a histone dimer from one side or the other, see Ramachandran et al. in the block resources document).
What do the differences between signal magnitudes (reflected by the y-axis) mean?
There are more positioned +1 nucleosomes then there are sub-nucleosomal intermediates, which can be rationalized by the fact that only a subset of genes are on / being transcribed in the cells at the time of the experiment.
Submit
Be sure to click the “Render” button to render the HTML output. Then paste the URL of this Posit Cloud project into the problem set on Canvas.